
サボテンは基本的にCAM型光合成を行うといわれています。
CAM型光合成の詳細な説明は省きますが、一般の植物が昼間、気孔を開き空気中の二酸化炭素を吸収し、日光のエネルギーを使ってデンプンなどを合成するのに対し(C3型光合成)、CAM 型光合成は、夜間気孔を開いて二酸化炭素を取り込み、主にリンゴ酸を合成し、昼間になると気孔を閉じて、夜間に合成されたリンゴ酸から二酸化炭素を再び発生させて、デンプンなどを合成するものです。
昼間に気孔を閉じている分、蒸散が抑えることができ、乾燥に強いのです。たしか。
・・・
それでは「葉っぱのついている原始的なサボテン」もCAM型光合成なのでしょうか。
葉っぱのついているサボテンの代表といえば、接ぎ木にもよく使われている「杢キリン」や「キリンウチワ(麒麟団扇)」でしょう。
というわけで杢キリンです。
接ぎ木の際は、枝を鉛筆状に削ってサボテンにぶっ刺すようです。台木としてはキリンウチワよりも成長が早いらしいですが、横方向に暴れやすいので扱いにくいとのことです(カクタス広瀬様談)。

キリンウチワです。
杢キリンと較べて茎が柔らかく緑色ですかね。
接ぎ木の際は、茎をカミソリ等でスパっと横に切って、実生苗などをぴょこんと置くようです。

実は見た目は似たようなこの2種の葉っぱサボテンですが、実は亜科がそもそも違います。
杢キリンはPereskia aculeataで「木の葉サボテン亜科(Pereskioideae)」。
キリンウチワはPereskiopsis velutinaで「団扇サボテン亜科(Opuntioideae)。
というわけで、この2つの亜科、見た目はサボテンには見えませんが、光合成についてはどうなっているのか、論文を読んでみます。
・・・アブストだけですけど。
※大学時代の一番頭のよかった時にTOEIC390点を叩きだした小生が訳しましたので、誤訳があったらごめんなさいです。
Leaf and Stem CO₂ Uptake in the Three Subfamilies of the Cactaceae, Park S. Nobel and Terry L. Hartsock, Plant Physiology, Vol. 80, No. 4 (Apr., 1986), pp. 913-917
Net CO2 uptake over 24-hour periods was examined for the leaves and
for the stems of 11 species of cacti representing all three subfamilies.
For Pereskia aculeata, Pereskia grandifolia, and Maihuenia poeppigli
(subfamily Pereskioideae), all the net shoot CO2 uptake was by the leaves
and during the daytime. In contrast, for the leafless species Carnegiea
gigantea, Ferocactus acanthodes, Coryphantha vivipara, and Mammillaria
dioica (subfamily Cactoideae), all the shoot net CO2 uptake was by
the stems and at night. Similarly, for leafless Opuntia ficus-indica
(subfamily Opuntioideae), all net CO2 uptake occurred at night. For leafy
members of the Opuntioideae (Pereskiopsis porteri, Quiabentia chacoensis,
Austrocylindropuntia subulata), at least 88% of the shoot CO2 uptake
over 24 hours was by the leaves and some CO2 uptake occurred at night.
Leaves responded to the instantaneous level of photosynthetically active
radiation (PAR) during the daytime, as occurs for C3 plants, whereas
nocturnal CO2 uptake by stems of 0. ficus-indica and F. acanthodes
responded to the total daily PAR, as occurs for Crassulacean acid
metabolism (CAM) plants. Thus, under the well-watered conditions
employed, the Pereskioideae behaved as C3 plants, the Cactoideae behaved
as CAM plants, and the Opuntioidea exhibited characteristics of
both pathways.
—日本語訳—3つのすべての亜科を表すサボテンの11種における葉および茎について、24時間にわたる二酸化炭素の取り込み純量が調査されました。Pereskia aculeata、Pereskia grandifolia、Maihuenia poeppigli(木葉サボテン亜科)については、シュートの二酸化炭素のすべての取り込みは、葉によって日中に行われました。対照的に、Carnegiea gigantea、Ferocactus acanthodes、Coryphantha vivipara、Mammillaria dioica(柱状サボテン亜科)については、シュートの二酸化炭素のすべての取り込みは、茎によって夜に起きました。同様に、葉のないOpuntia ficus-indica(団扇サボテン亜科)については、二酸化炭素の取り込みは夜に行われました。団扇サボテン亜科における葉のあるもの(Pereskiopsis porteri、Quiabentia chacoensis、Austrocylindropuntia subulata)については、24時間にわたるシュートの二酸化炭素取り込み量の少なくとも88%は葉によって行われ、いくつかは夜に起きました。葉は日中における光合成有効放射(PAR)の瞬時レベルに応答しましたが(C3型植物のために起こるように)、その一方で0. ficus-indica およびF. acanthodesの茎による夜間の二酸化炭素取り込みはトータルの日中の光合成有効放射に応答しました(CAM型植物のために起こるように)。このように、水の多い状況の下では、木葉サボテン亜科はC3型植物として振る舞い、柱状サボテン亜科はCAM型植物として、そして団扇サボテン亜科は両方の方式の特徴を示します。
どうやら、杢キリン(木の葉サボテン亜科)はC3型光合成で、キリンウチワ(ウチワサボテン亜科)はC3とCAMの両方のようです。なかなか興味深いですね。
もう一つ読んでみましょう。
Craig E. Martin1 and Robert S. Wallace,2000,Photosynthetic Pathway Variation in Leafy Members of Two Subfamilies of the Cactaceae,Int. J . Plant Sci. 161(4):639-650.
Patterns of 24‐h CO2 exchange and diel fluctuations in tissue acid concentrations were measured in leafy and leafless shoots of 10 species in the Pereskioideae and eight species in the Opuntioideae (Cactaceae). The species were selected to represent a range of phylogenetic histories. Leafy shoots of all species in the Pereskioideae exhibited C3 patterns of gas exchange, and net CO2 exchange of leafless stems in all but one species was negative during the day and night. Although nighttime CO2 uptake was not observed in shoots or stems of any of the pereskioid taxa, tissue acidity increased at night to a small degree in leaves of six species and stems of five species, indicative of low levels of CAM‐cycling. In contrast, in leafy shoots of nearly all species in the Opuntioideae, CO2 uptake occurred during the day and the night. Gas‐exchange rates were typically greater during the day. As is typical of CAM, nighttime maximal water use efficiency often greatly exceeded daytime values. Tissue malic acid concentrations increased overnight in leaves and stems of all eight opuntioid species. Examination of the data from a phylogenetic perspective illustrates evidence of low levels of CAM scattered among the primarily C3 members of the more ancestral Pereskioideae. Furthermore, such consideration of the taxa in the more derived Opuntioideae (comparing the genera from most ancestral to most derived, that is,Austrocylindropuntia → Quiabentia → Pereskiopsis → Cylindropuntia) revealed that CAM became increasingly less important in the leaves of the various taxa, whereas this water‐conservative pathway of photosynthesis became increasingly more important in the stems. The results of this study indicate that members of the Pereskioideae should be restricted to moister habitats or must restrict the timing of growth to wet seasons, whereas the observed combinations of the C3 and CAM pathways in the opuntioid taxa should prove beneficial in conserving water in the sporadically arid tropical and subtropical habitats of these plants.
—日本語訳—
24時間の二酸化炭素交換のパターンと細胞組織内酸濃度の日周変動が、木の葉サボテン亜科の10種、団扇サボテン亜科の8種の葉有りおよび葉無しのシュートについて調査されました。その種達は、系統発生学的な歴史の範囲を表すために選ばれました。木の葉サボテン亜科におけるすべての種の葉有りのシュートは、ガス交換のC3パターンを示し、そして日夜間における、一種を除いた葉無しの茎の純二酸化炭素交換量はマイナスでした。どんなペレスキオイドの分類群のシュートおよび茎においても、夜間の二酸化炭素の取り込みは観測されなかったにもかかわらず、細胞内酸度は夜に6種の葉および5種の茎において、少し程度増加しました(CAM型サイクルの低いレベルを示します)。対照的に、団扇サボテン亜科のほとんどすべての葉有りのシュートについては、二酸化炭素の取り込みは日夜間でおきました。ガス交換の割合は、概して日中のほうが高かったです。CAM型の典型であるように、夜間の最大の水利用効率は、しばしば日中の値を超えました。細胞組織内のリンゴ酸濃度は8のすべてオプンチオイド種の葉と茎において、夜に渡り増加しました。系統発生学的観点からのデータの検討は、より多くの先祖の木の葉サボテン亜科の主にC3のメンバーたちの間に、散りばめられたCAM型の低いレベルを説明します。 さらに、より多く派生した団扇サボテン亜科(殆どの先祖から殆どの由来された属との比較、すなわち、アウストロキリンドロプンチア→クイアベンチア→ペレスキオプシス→キリンドロプンチア)における、このような分類学的検討は、CAM型がさまざまな分類学的な葉において、だんだん重要でなくなったことに反して、光合成のこの保水過程は茎においてだんだん重要ようになってきたことを明らかにします。この研究の結果は、木の葉サボテン亜科のメンバーは湿った生息場所に制限するべき、もしくは成長のタイミングを、湿気のある季節に制限しなければならないのに反して、オプンチオイド分類学群におけるC3型とCAM型の観測されたコンビネーションは、これらの植物の、単発的に乾燥した熱帯および亜熱帯の生息地での、水を保存することに有益であることを証明するべきものであると指し示します。
忠実に訳したせいで、意味不明な部分もあるかと思いますがご了承ください。
また、ペレスキオプシス(ウチワサボテン亜科)の「C3型光合成+CAM型光合成」という不思議な光合成方式について、わかりやすいデータが以下の論文にありましたので、グラフにしてみました。
Nobel PS, Hartsock TL (1987) Drought-induced shifts in daily CO2 uptake patterns for leafy cacti. Physiologia Plantarum 70, 114-8.
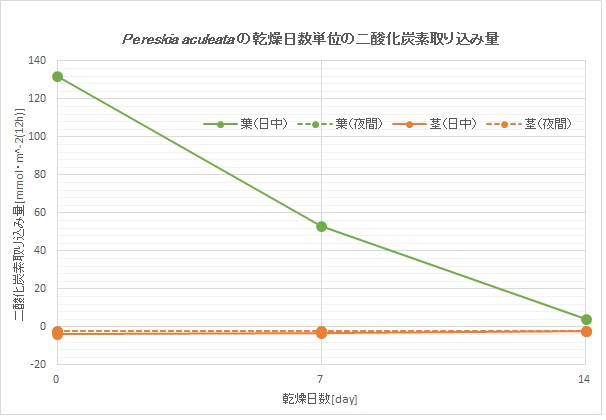
下のグラフはペレスキア(木の葉サボテン亜科)のとある種の、乾燥に対する二酸化炭素取り込み量の関係です。
水をあげないでおくと、二酸化炭素取り込み量(すべてが日中の葉によるもの)はどんどん減っていきます。C3型光合成の典型的なものだと思います。
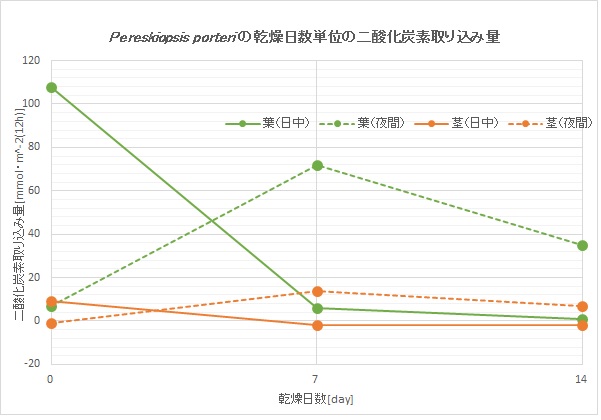
下のグラフはペレスキオプシス(ウチワサボテン亜科)のとある種のものです。
水をあげないと、日中の葉による二酸化炭素取り込み量はペイレスキアのようにどんどん減っていきますが、それと同時に乾燥が始まると夜間に葉と茎による二酸化炭素取り込み量が増えていきます(CAM型光合成)。
というわけでまとめです。
<杢キリン>
- 光合成はC3型光合成
- 光合成は水が多い時はC3型光合成、乾燥するとCAM型光合成。
というわけで、サボテンの成長が遅いのはCAM型光合成のためであると言われていますので、これら2つに接ぎ木するとC3型光合成の力を借りることができ、成長が早いということですね。たぶん。
そんで、接ぎ木した時は、水をジャバジャバあげて乾燥させないほうがよく、光合成の主体となる葉っぱを大事にしなければならないということです。
というわけで勉強になりました!

